
SEEFOR 17(1): 26008
Article ID: 26008
DOI: https://doi.org/10.15177/seefor.26-008
ORIGINAL SCIENTIFIC PAPER
Community Structure, Diversity, and Population Dynamics of Black Poplar Leaf Miners and Galling Aphids in the Urban Environment
Jovan Dobrosavljević1,*, Marjanović Marija2, Čedomir Marković1
Addresses:
(1) University of Belgrade, Faculty of Forestry, Department of Forestry, Kneza Višeslava 1, RS-11030 Belgrade, Serbia;
(2) University of Belgrade, Faculty of Forestry, Department of Landscape Architecture and Horticulture, Kneza Višeslava 1, RS-11030 Belgrade, Serbia
Citation: Dobrosavljević J, Marjanović M, Marković Č, 2026. Community Structure, Diversity, and Population Dynamics of Black Poplar Leaf Miners and Galling Aphids in the Urban Environment. South-east Eur for 17(1): 26008. https://doi.org/10.15177/seefor.26-008 .
Received: 19 Jan 2026; Revised: 27 Mar 2026; Accepted: 29 Apr 2026; Published online: 27 Jun 2026
Cited by: Google Scholar
Abstract
Trees in the urban environment, such as the black poplar (Populus nigra L.), are strongly affected by environmental changes, which can even lead to their decline. Since the development of large cities on the river banks, black poplar is losing its habitat, but is also threatened by factors such as pollution, climate change, and altered habitat characteristics. These changes affect the insects feeding on black poplar, especially the endophagous species, which are in close contact with the host through their entire development, such as the leaf miners (LMs) and galling aphids (GAs). That is why we examined the community structure, diversity, and population dynamics of black poplar LMs and GAs. The study was conducted in Belgrade, Serbia, on five urban sites differing in vegetation characteristics. Sampling was conducted once per month in the period May-September, in 2016 and 2017. Thirteen species of LMs and seven GAs were identified on the studied trees. The seasonal and site differences affected the LM and GA communities, showing their different adaptation strategies for coexistence on the same host, while the effect of the study year was modest. Greater GAs species richness and LMs diversity were recorded on sites with higher participation of canopy and green area cover. The results highlight the importance of urban green structure for sustaining specialized herbivorous insects and suggest that LMs and GAs on black poplar can serve as sensitive indicators of ecological quality in urban environments.
Keywords: abundance; green areas; canopy cover; Populus nigra L.; seasonal dynamics; species richness
INTRODUCTION
With increasing urbanization, many tree species are disappearing from the cities or are being suppressed to the city edges (Helen et al. 2019, Korányi and Markó 2022, Łukaszkiewicz et al. 2024, Xie et al. 2024). One such species is the black poplar (Populus nigra L.), a key tree species of floodplain and riverbank ecosystems in the basins of major European rivers (Lefèvre et al. 2001, de Rigo et al. 2016, Łukaszkiewicz et al. 2024). Black poplars are ecologically significant as they stabilize and remedy the soil in sensitive habitats, sustain great biodiversity, and improve air quality due to their rapid growth and large leaf area (Gilman and Watson 1994, Lefèvre et al. 2001, Šiler et al. 2014, de Rigo et al. 2016, Levei et al. 2021, Łukaszkiewicz et al. 2024). However, several of its traits are considered a drawback for urban environments: its brittle branches break easily and can pose safety risks, its strong roots can damage infrastructure, and its abundant pollen and fluffy seeds can cause discomfort to people (Gilman and Watson 1994, Costello et al. 1997, Minić et al. 2020, Costache et al. 2021, Shams et al. 2021, Łukaszkiewicz et al. 2024).
Besides the direct effect of urbanization on the decline of black poplar from the urban environment, the trees in the cities can be affected in many ways (Rickman and Connor 2003, Dobrosavljević et al. 2020, 2023, Korányi and Markó 2022, Percival 2023, Franceschi et al. 2023, Gao et al. 2023). Elevated temperatures and air pollution in the urban environment can cause significant stress, which is further increased by altered soil properties and groundwater regimes (Percival 2023, Franceschi et al. 2023, Gao et al. 2023). These, and other environmental disturbances, negatively affect organisms trophically connected to the black poplar trees (Southwood 1961, Schowalter 1995, Dobrosavljević et al. 2023). Among these organisms, insects are the most numerous and significant (Southwood 1961, Lefèvre et al. 2001, Vanden Broeck 2003). The majority of them are herbivorous species, which are highly sensitive to environmental change (Raupp et al. 2010, Valdés-Correcher et al. 2022, Dobrosavljević et al. 2023). They are affected both directly and indirectly, through the host plants (Schowalter 2006, Jones and Leather 2012, Sallé et al. 2021). The most sensitive are the species that cannot migrate far from their hosts, and those that develop in close contact with their hosts, such as the leaf miners and gall-inducing insects (Moir et al. 2014, Dobrosavljević et al. 2020, 2023, Sallé et al. 2021, Valdés-Correcher et al. 2022). They respond to environmental changes by altering their population spatial distribution and seasonal dynamics (Sunil et al. 2023, Boggs 2024, Ojija et al. 2025). LMs and GAs are diverse insect groups with numerous species that are taxonomically well studied in the study area and globally distributed across a wide range of habitats (Hałaj and Osiadacz 2013, Dobrosavljević et al. 2017, Ellis 2020, Dobrosavljević and Marković 2024).
As many European capitals are located near big rivers, the black poplar is becoming increasingly endangered (Lefèvre et al. 2001, de Rigo et al. 2016). Its status is further threatened by the practice of conversion of wetlands to agricultural land, which is now largely prohibited across Europe (Ramsar Convention Secretariat 2016). To determine how the insect community inhabiting black poplar behaves in an urban setting, we investigated the community structure, diversity, and population dynamics of LMs and GAs. The study aimed to determine the effect of (1) interannual variation, (2) seasonal dynamics, and (3) site characteristics on the LMs and GAs abundance, species richness, and diversity in the urban environment.
MATERIALS AND METHODS
Study Design and Sampling
The study was conducted in 2016 and 2017 on five sites in Belgrade: Ada Ciganlija (44°47'09"N, 20°24'41"E), Dorćol (44°49'49"N, 20°27'29"E), New Belgrade block 69 (44°48'11"N, 20°25'14"E), New Belgrade Studenjak (44°49'32"N, 20°24'00"E), and Rakovica (44°46'13"N, 20°26'23"E). On each site, five trees measuring at least 30 cm in diameter and at least 10 m in height were selected. Once per month in the period May-September, five 50‑100 cm long lateral branches with leaves were collected from each tree to a height of about 4 m (5 branches · 5 trees · 5 sites = 125 branches per session or 25 branches per site).
All samples were brought to the Entomological laboratory of the Faculty of Forestry, University of Belgrade, where the total number of leaves was counted. Leaves with mines and galls were separated and individually analyzed. The presence of live larvae or pupae within mines and galls was recorded. However, not all mines and galls contained live individuals due to factors such as prior emergence, mortality, or parasitism.
Species identification was primarily based on the characteristics of mines and galls. If immature stages (eggs, larvae, pupae, larval cases, and cocoons) were present in the analyzed material, they were examined and used for morphological identification. To increase the reliability of the identification and to confirm the species’ identity, adults were reared from a part of the collected larvae and pupae (Hering 1957, Blackman and Eastop 1994, Hałaj and Osiadacz 2013, Doorenweerd et al. 2014, Haarder and Liston 2018, Laštůvka et al. 2018, Langmaid 2019, Lepiforum 2019, Liston et al. 2019, Ellis 2020, Babichev and Kirichenko 2020).
Site Characteristics
Belgrade, the capital of Serbia, was selected as the study area because it represents a large urban centre situated at the confluence of two major rivers, the Sava and the Danube. This geographic setting, combined with diverse levels of urbanization and green space distribution, makes Belgrade an ideal environment for studying how urban conditions and proximity to riverbanks influence insect communities associated with black poplar. The trees at the sampling sites were all distributed in groups of up to 10 trees. All of the studied sites were located in urban areas, New Belgrade and Dorćol in heavily urbanized areas, while Ada Ciganlija and Rakovica in areas with slightly less intense urbanization. Sites New Belgrade block 69, Dorćol, and Ada Ciganlija were located in the immediate vicinity of the Sava and Danube rivers. At the same time, sites New Belgrade, Studenjak, and Rakovica were more than 500 meters from the large river banks. At the sampling sites, 200 m diameter buffer zones were selected. In each buffer zone, the green spaces (grasslands, parks, dirt trails, etc.), spaces with no greenery (impervious areas, bodies of water, railroad tracks, etc.), as well as the total area covered by tree canopies were measured. The aerial images used for the analysis were obtained from Google Earth Pro 7.3.6.10201, from the images made in 2018 (Google Earth Pro (Desktop), 2025). ImageJ 1.53e (Schneider et al. 2012) was used for all the area measurements and creation of the buffer zone circles.
Characterisation of the Leaf Miner (LM) and Galling Aphid (GA) Community Analyzed
LM and GA communities were quantified by the following parameters at the level of a single tree (Dobrosavljević et al. 2020, 2023):
The abundance of individual species (Ai) is calculated
as the number of mines per 100 leaves:
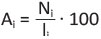 (1)
(1)
where ni is the number of mines of one particular species found in one sample, and li is the number of leaves per sample.
The total abundance of (Ab) is calculated as the sum
of all individual species’ abundance.
Species richness (S) (number of species identified).
Shannon’s index of diversity (H'), calculated as:
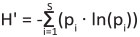 (2)
(2)
where S is the Species richness and pi is the proportion of individuals belonging to the i-th species in the dataset (Shannon 1948).
Dominant species were defined as the subset of the most abundant species that cumulatively accounted for 90% of the total LMs or GAs abundance in a given year. The 90% threshold was selected to capture the core community while excluding rare species (Magurran 2004).
Statistical Analysis
As the Kolmogorov–Smirnov test indicated that the data distribution does not fit any of the standardized distributions, nonparametric methods were used for the analysis. The differences in the LM and GA abundance, species richness, and diversity between the months in the vegetative season and locations were determined by the Kruskal–Wallis test, while the differences between the studied years were determined by the Mann–Whitney U test. Only dominant species were included in the individual species analysis. When the Kruskal–Wallis test highlighted significance, the Mann-Whitney U test was used as a post hoc test to conduct pairwise analysis. Linear regression was used to determine how the analyzed parameters relate to the green spaces and canopy cover of the investigated sites. Data were analyzed at a significance level of p < 0.05. The diversity index was calculated in PAST 5.2.2. (Hammer et al. 2001). All of the statistical analyses were conducted in Statistica 12.0 (StatSoft 2014).
RESULTS
Thirteen species of LMs and seven species of GAs were identified on the leaves of the studied trees (Table 1). The most abundant LM species were Phyllocnistis unipunctella (Stephens, 1834), Ph. extrematrix Martynova, 1955, and Stigmella trimaculella (Haworth, 1828), while the most abundant GAs were Pemphigus spyrothecae Passerini, 1860 and Thecabius lysimachiae Börner, 1916.
Dominant LM species in 2016 were by their participation in the total abundance Ph. unipunctella (27.5%), Ph. extrematrix (27.1%), S. trimaculella (19.9%), and A. populi (13.1%), while in 2017 the community was dominated by Ph. unipunctella (40.2%), L. sinuella (20.2%), P. populifoliella (10.8%), S. trimaculella (7.9%), Ph. extrematrix (5.4%), and F. hortulana (4.1%). Dominant GA species in 2016 were, by their participation in the total, P. spyrothecae (50.7%), T. lysimachiae (19.9%), T. affinis (13.4%), and P. populinigrae (5.7%), while in 2017 the community was dominated by T. lysimachiae (46.7%), P. spyrothecae (21%), T. affinis (12.7%), and P. populinigrae (7.9%).
LM abundance differed significantly between the investigated years for all the dominant species except Ph. extrematrix (Table 1). Only half of the dominant GA species were affected by the interannual variation (T. affinis and T. lysimachiae) (Table 1). The dominant LM species were affected by the differences between the months in the vegetative season, both in 2016 and 2017 (Figure 2a,b, Table A1), while the GAs species showed a different pattern, as only one species was significantly affected by the seasonality, only in 2016 (Figure 2c,d, Table A1). Site characteristics significantly affected all the dominant LM and GA species in both of the analyzed years (Table 2, Table A1).
The total LM and GA abundance did not differ significantly between the investigated years (Z = -1.258,
p = 0.208; Z = 0.839, p = 0.401). LM abundance was highest in August, while GA abundance remained relatively consistent throughout the season, with the highest values noted in May (Figure 3a,c). Month of the vegetative season as well as site characteristics affected both LMs
(H = 108.542, p < 0.01; H = 22.764, p < 0.01) and GAs abundance (H = 241.001, p < 0.01; H = 12.140, p = 0.016) (Figure 3, Table A2, Table A3).
LM species richness and diversity did not differ significantly between the two years (Z = - 1.097,
p = 0.265; Z = 0.868, p = 0.385). GA species richness differed significantly between the two years (Z = 2.002, p = 0.045), while the diversity did not (Z = 1.551, p = 0.060). Further down, the characteristics of the community that did not vary between years were analyzed using pooled data, whereas those that showed yearly variation were analyzed separately for each year.
The total LM species richness was affected by both the study site and seasonality (H = 13.923, p = 0.008;
H = 76.930, p < 0.001) (Figure 4, Table A4), while the diversity was affected only by seasonality (H = 20.745, p = 0.004) (Figure 5a,b, Table A5). GA species richness was affected by the locality both in 2016 and 2017 (H = 28.381, p < 0.001; H = 75.649, p < 0.001) (Figure 6b,d, Table A4), while it was not affected by seasonality in neither in 2016, nor in 2017 (H = 1.783, p = 0.776; H = 5.467, p = 0.243) (Figure 6a,c, Table A4). The total GA diversity was only affected by the study site (H = 86.775, p < 0.001) (Figure 5c,d, Table A5).
Green areas in the 200 m radius buffer zone comprised, on average, 53.4 ± 14.26% of the total area, while the tree canopies covered, on average, 29.4 ± 13.31% of the total area. The site with the greatest green area participastion was Rakovica (79%), while Dorćol had the lowest (35%), mainly because the water area comprised a large part of the buffer zone (31%). Tree canopy cover was greatest at NBG Studenjak (45%), and lowest at NBG block 69 (9%) (Figure 1, Table 3).
Although the number of investigated sites was not great enough for a detailed insight into the effect of their characteristics on the studied insect communities, we determined that greater green area and canopy cover positively affected GA abundance and species richness, LM diversity, while the green area on the studied sites affected GA species richness and LM diversity (Table 4).
DISCUSSION
The results of this study highlight that different group of black poplar endophagous insects - leaf miners (LMs) and galling aphids (GAs) - respond differently to changes in environmental and temporal factors in the urban environment. Among the analyzed drivers, interannual variation had the lowest influence on the overall community patterns, whereas microclimatic and habitat-related factors played a more prominent role, consistent with previous studies on urban insect assemblages (Korányi and Markó 2022, Dobrosavljević et al. 2023).
Interannual differences significantly affected GA species richness, but their impact on community-level abundance remained limited. However, the differences between the studied years had a significant effect on the abundance of most of the dominant LM species, such as S. trimaculella and Ph. unipunctella. These species were abundant in both years, but the environmental differences between the two studied years affected them in opposite ways. While the abundance of S. trimaculella declined, Ph. unipunctella increased its abundance significantly, which can be a consequence of utilizing the increased available resources, such as leaf availability and improved leaf quality, according to Balaji et al. 2024. This highlights the importance of individual species biology for adaptation to even subtle differences in environmental conditions, and stressors such as temperature or humidity, as well as diversity of surrounding plants, and other factors (Connor and Taverner 1997, Rickman and Connor 2003, Fernandes et al. 2004, Rodrıguez-Castañeda et al. 2017, Barantal et al. 2019, Dobrosavljević et al. 2023, Chen et al. 2023). These differences show that even though the individual species were affected by the interannual variation, the overall community abundance remained stable, suggesting a compensatory response among dominant species that maintained overall community stability. On the other side, the interannual effect was not that significant for the GA community, as only half of the dominant species were affected. P. spyrothecae, the most abundant GA species in this study, likely maintained its population levels due to biological traits like the protective features of the gall, as noted in previous studies (Blackman and Eastop 1994, Alton 1999, Hałaj and Osiadacz 2013, Kot and Kmieć 2020). The effect of interannual variation on the LM and GA communities is likely to become more pronounced under climate change, as growing variability in temperature, humidity, and precipitation can directly influence insect development, survival, and phenology, as well as indirectly alter host plant quality and defenses (Bale et al., 2002, Parmesan, 2006, Pincebourde et al. 2012, Robinet and Roques, 2010). Additionally, the increasing frequency of extreme climatic events may further amplify year-to-year fluctuations in insect populations, particularly in urban environments where microclimatic conditions are already highly heterogeneous (Oke 1982, Meineke et al. 2013, Korányi and Markó 2022). The importance of the interannual variation, due to climate change, as many environmental factors, such as the temperature, humidity, and precipitation patterns, will change. That is why long-term monitoring of the insect-plant interactions, especially in the urban environment, is necessary.
The LM and GA communities reacted differently to seasonal variation. The LMs' abundance, species richness, and diversity peaked in the middle of the studied period (July and August), similar to previous studies conducted in both rural and urban environments (Southwood et al. 2004, Dobrosavljević et al. 2020, Marković et al. 2021b). This trend also occurs in other insect species such as coccinellids and LM communities on other host plants (Southwood et al. 2004, Marković et al. 2018). This is a consequence of the better leaf quality as food for the LMs and interguild competition in the earlier part of the year (Connor and Taverner 1997, Southwood et al. 2004). Another reason why the LM community peaked in the middle of the studied period is that most of the dominant LM species are multivoltine, and their second generation, which feeds during July and August, is more abundant than the first generation (Hering 1951, Ellis 2020). The only dominant species that did not peak during this period was Fenusella hortulana, as it develops only one generation in early spring (Hering 1951, Ellis 2020, Dobrosavljević et al. 2023). In contrast, the GA community characteristics peaked much earlier in the vegetative season, in the period May-June. One of the reasons for this lies in the behaviour of the most abundant dominant species, P. spyrothecae, which tends to form galls on the petioles of young, developing leaves (Blackman and Eastop 1994, Alton 1999, Hałaj and Osiadacz 2013, Kot and Kmieć 2020). Their contrasting seasonal dynamics suggest temporal partitioning between the two endophagous herbivore guilds, which may facilitate their coexistence on the same host trees by reducing potential overlap in resource use. Besides the vegetative season, the studied guilds, especially the GA community, which was less affected by the seasonal variation, were affected by habitat characteristics. We determined that the analyzed habitat characteristics (percentage of green area and canopy cover on the 200 m buffer zones) had a significant effect on some of the investigated insect community characteristics, although not with a strong correlation coefficient (GA abundance and species richness, and LM diversity), similar to other studies (Knuff et al. 2020, Fenoglio et al. 2023, Matilda Collins et al. 2024, Abrego and Medianero 2025, Sinclair et al. 2025). This highlights the importance of urban greenery, especially of the urban trees, for diversity conservation. Even though the investigated factors influenced the LM and GA communities, there are also many other factors such as landscape, microclimate, host trees characteristics, predator and parasitoid community characteristics, etc. which can also affect insect communities in the urban environment (Rickman and Connor 2003, Baraniak et al. 2004, Fenoglio et al. 2009, Bairstow et al. 2010, Peralta et al. 2011, Jones and Leather 2012, Cebeci et al. 2018, Milanović et al. 2020, Dobrosavljević et al. 2020, 2023, 2025, Marković et al. 2021a, Valdés-Correcher et al. 2022, Matilda Collins et al. 2024, Horák et al. 2024). That is why more detailed studies, which can give insights into the influence of other individual environmental factors on the herbivorous insect behaviour in urban environments, need to be conducted. All this suggests that habitat characteristics and seasonal variation have a stronger effect on the investigated insect guilds than interannual variation, similar to studies on plant-insect interactions in similar climate conditions (Southwood et al. 2004, Korányi and Markó 2022).
Although the LMs and GAs behave similarly (both feed endophagously on the same host plant), they are differently affected by the environmental differences. This is even more pronounced on the level of individual species, where, i.e. the abundance of the miner P. unipunctella varies significantly between years and sites, reflecting its sensitivity to microhabitat conditions, while the dominant aphid species P. spyrothecae shows a more stable dynamics due to the differences in their biology (Hering 1951, Alton 1999, Ellis 2020, Kot and Kmieć 2020). Although LMs and GAs comprise a significant component of black poplar insect diversity, the influence of environmental changes caused by urbanization and climate change on other insects needs to be addressed, as black poplar is a host for more than 100 insect species (Southwood 1961). Conversely, insects can cause significant damage to trees, especially when they are young and sensitive (Marković et al. 2024), which is why the significance of these insect species as pests should be reviewed. Even after identifying significant pests, their management in the urban environment is challenging, as chemical insecticides, which could be used in their natural habitat, can pose risks for the already fragile environment, and even for people in the cities (Md Meftaul et al. 2020). This highlights the importance of constant monitoring of insect populations in the cities, and their integrated pest management (IPM) approaches (Dreistadt and Flint 1995, Dhang 2014, Tabassum et al. 2024, Milanović et al. 2025). Integrating such strategies into urban forestry planning can help balance tree protection, biodiversity conservation, and ecosystem health, thereby supporting the black poplar and its insect communities.
CONCLUSIONS
This study showed that leaf miners (LMs) and galling aphids (GAs) on black poplar in urban habitats react differently to temporal and environmental changes. The interannual variation had a modest influence, affecting only GA species richness and a few dominant LM species. Seasonal changes and habitat characteristics influenced their abundance and diversity much more significantly. While GAs abundance remained relatively consistent throughout the season, with the highest values noted in May, LMs' notably peaked mid-summer, reflecting the differences in their life cycles and survival strategies. Such differences may facilitate their coexistence on the same host trees by reducing overlap in resource use, rather than implying direct competitive interactions. Habitat characteristics, particularly the proportion of green areas and canopy cover, were important drivers of insect community structure, highlighting the role of urban greenery in supporting biodiversity. At the species level, responses were highly variable, emphasizing the importance of species-specific traits in shaping population dynamics under urban conditions. Given the ecological importance and ongoing decline of black poplar in cities, these findings underline the need for its conservation and for the implementation of sustainable, ecologically based pest management strategies. Overall, the results contribute to a better understanding of plant–insect interactions in urban ecosystems and provide a basis for integrating biodiversity conservation with urban forestry planning.
Author Contributions
JD, ČM and MM conceived and designed the research, JD, ČM and MM carried out the field measurements, JD performed laboratory analysis, JD processed the data and performed the statistical analysis, JD and ČM secured the research funding, supervised the research and helped to draft the manuscript, JD and ČM wrote the manuscript.
Funding
This work was supported by the Serbian Ministry of Education, Science and Technological Development, grant number 451-03-137/2025-03/200169, for financing scientific research at the University of Belgrade, Faculty of Forestry.
Conflicts of Interest
The authors declare no conflicts of interest.
Apendix
REFERENCES
Abrego J, Medianero E, 2025. The diversity and composition of insect communities in urban forest fragments near Panama City. Biology (Basel) 14(6): 721. https://doi.org/10.3390/BIOLOGY14060721.
Alton KL, 1999. The biology of Pemphigus spyrothecae galls on poplar leaves (PhD thesis). University of Nottingham, Nottingham, England, 259 p.
Babichev N, KirichenkoN, 2020. Diversity and distribution of gall-forming aphids of the genus Pemphigus (Sternorrhyncha: Aphididae, Pemphigini) in Eastern Siberia. J Asia Pac Biodivers 13(3): 339–348. https://doi.org/10.1016/j.japb.2020.07.002.
Bairstow KA, Clarke KL, McGeoch MA, Andrew NR, 2010. Leaf miner and plant galler species richness on Acacia: Relative importance of plant traits and climate. Oecologia 163: 437–448. https://doi.org/10.1007/s00442-010-1606-4.
Balaji BN, Kumaraswamy S, Pavithra V, 2024. Co-occurrence and abundance of two leafminer species, Phthorimaea absoluta (Lepidoptera: Gelechiidae) and Liriomyza trifolii (Diptera: Agromyzidae) on tomato. J Entomol Res 48(4): 574–577. https://doi.org/10.5958/0974-4576.2024.00111.7.
Bale JS, Masters GJ, Hodkinson ID, Awmack C, Bezemer TM, Brown VK, Butterfield J, Buse A, Coulson JC, Farrar J, Good JEG, Harrington R, Hartley S, Jones TH, Lindroth RL, Press MC, Symrnioudis I, Watt AD, Whittaker JB, 2002. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob Change Biol 8(1): 1–16. https://doi.org/10.1046/j.1365-2486.2002.00451.x.
Baraniak E, Walczak U, Tryjanowski P, Zduniak P, 2004. Effect of distance between host trees and leaf litter removal on population density of Cameraria ohridella Deschka & Dimic, 1986 (Lepidoptera, Gracillariidae) - Pest of chestnut (Aesculus sp.) trees. Pol J Ecol 52(4): 569–574.
Barantal S, Castagneyrol B, Durka W, Iason G, Morath S, Koricheva J, 2019. Contrasting effects of tree species and genetic diversity on the leaf-miner communities associated with silver birch. Oecologia 189: 687–697. https://doi.org/10.1007/s00442-019-04351-x.
Blackman RL, Eastop VF, 1994. Aphids on the world’s trees: an identification and information guide. CAB International, Wallingford, Oxfordshire, UK, 1024 p.
Boggs CL, 2024. Changes in insect population dynamics due to climate change. In: González-Tokman D, Dáttilo W (Eds) Effects of Climate Change on Insects: Physiological, Evolutionary, and Ecological Responses. Oxford Academic, Oxford, UK, pp 157–177. https://doi.org/10.1093/OSO/9780192864161.003.0009.
Cebeci HH, Markovic C, Grabenweger G, Ayberk H, Dobrosavljevic J, Goltas M, Stojanovic A, Ale A, Bacchetta C, Cazenave J, 2018. Preliminary notes on pupal parasitism rates of the horse chestnut leafminer, Cameraria ohridella (Lepidoptera Gracillariidae) in Belgrade and Istanbul. Fresenius Environ Bull 27(10): 7122–7124.
Chen X, Zhong M, Cui L, Xu J, Dai X, Liu X, 2023. Elevational Pattern of Leaf Mine Diversity on Quercus variabilis Blume at Baotianman, Henan, China. Insects 14(1): 7. https://doi.org/10.3390/insects14010007.
Connor EF, Taverner MP, 1997. The Evolution and Adaptive Significance of the Leaf-Mining Habit. Oikos 79(1): 6–25. https://doi.org/10.2307/3546085.
Collins CM, Audusseau H, Hassall C, Keyghobadi N, Sinu PA, Saunders ME, 2024. Insect ecology and conservation in urban areas: An overview of knowledge and needs. Insect Conserv Divers 17(2): 169–181. https://doi.org/10.1111/ICAD.12733.
Costache A, Berghi ON, Cergan R, Dumitru M, Neagos A, Popa LG, Giurcaneanu C, Vrinceanu D, 2021. Respiratory allergies: Salicaceae sensitization (Review). Exp Ther Med 21(6): 609. https://doi.org/10.3892/ETM.2021.10041.
Costello LR, Elmore CL, Steinmaus S, 1997. Tree Root Response to Circling Root Barriers. AUF 23(6): 211–218. https://doi.org/10.48044/JAUF.1997.033.
De Rigo D, Enescu CM, Houston Durrant T, Caudullo G, 2016. Populus nigra in Europe: distribution, habitat, usage and threats, in: San- Miguel-Ayanz J, de Rigo D, Caudullo G, Houston Durrant T, Mauri A. (Eds.), European Atlas of Forest Tree Species. Publ. Off. EU, Luxembourg, pp. 136–137. https://doi.org/10.2760/776635.
Dhang P (Ed.), 2014. Urban insect pests: sustainable management strategies. CABI publishing, Wallingford, Oxfordshire, United Kingdom, pp 1-201. https://doi.org/10.1079/9781780642758.0000.
Dobrosavljević J, Kanjevac B, Marković Č, 2025. Microclimate Shifts and Leaf Miner Community Responses to Shelterwood Regeneration in Sessile Oak Forests. Forests 16(5): 739. https://doi.org/10.3390/f16050739.
Dobrosavljević J, Marković Č, 2024. First Findings of Deciduous Woody Plant Leaf Miners in Serbia. J Entomol Res Soc 26(2): 183–207. https://doi.org/10.51963/jers.v26i2.2498.
Dobrosavljević J, Marković Č, Bojić S, 2017. Overview of leaf miner fauna in Serbia. In: Kovačević D, (ed) Proceedings of the VIII International Agriculture Symposium “AGROSYM 2017”, Jahorina, Bosnia and Herzegovina, 05 - 08 October 2017. East Sarajevo University Faculty of Agriculture, East Sarajevo, Bosnia and Herzegovina, pp 1490–1498.
Dobrosavljević J, Marković Č, Marjanović M, 2023. The effect of urban–rural gradient on black poplar endophagous herbivorous insects. Arthropod Plant Interact 17: 341–350. https://doi.org/10.1007/s11829-023-09963-y.
Dobrosavljević J, Marković Č, Marjanović M, Milanović S, 2020. Pedunculate Oak Leaf Miners’ Community: Urban vs. Rural Habitat. Forests 11(12): 1300. https://doi.org/10.3390/f11121300.
Doorenweerd C, Van Haren MM, Schermer M, Pieterse S, Van Nieukerken EJ, 2014. A Linnaeus NGTM interactive key to the Lithocolletinae of North-West Europe aimed at accelerating the accumulation of reliable biodiversity data (Lepidoptera, Gracillariidae). Zookeys 422: 87–101. https://doi.org/10.3897/zookeys.422.7446.
Dreistadt SH, Flint ML, 1995. Landscape Pest Monitoring Methods and Training Managers to Use Them. AUF 21(1): 1–6. https://doi.org/10.48044/JAUF.1995.001.
Ellis WN, 2020. Leafminers and plant galls of Europe. Available online: http://www.bladmineerders.nl (12 December 2025).
Fenoglio MS, González E, Tavella J, Beccacece H, Moreno ML, Fabian D, Salvo A, Estallo EL, Calviño A, 2023. Native plants on experimental urban green roofs support higher community-level insect abundance than exotics. Urban For Urban Green 86: 128039. https://doi.org/10.1016/J.UFUG.2023.128039.
Fenoglio MS, Salvo A, Estallo EL, 2009. Effects of urbanisation on the parasitoid community of a leafminer. Acta Oecol 35(2): 318–326. https://doi.org/10.1016/j.actao.2008.12.001.
Fernandes GW, Caldeira Castro FM, Faria ML, Marques ESA, Barcelos Greco MK, 2004. Effects of Hygrothermal Stress, Plant Richness, and Architecture on Mining Insect Diversity. Biotropica 36: 240–247. https://doi.org/10.1111/J.1744-7429.2004.TB00315.X.
Franceschi E, Moser-Reischl A, Honold M, Rahman MA, Pretzsch H, Pauleit S, Rötzer T, 2023. Urban environment, drought events and climate change strongly affect the growth of common urban tree species in a temperate city. Urban For Urban Green 88: 128083. https://doi.org/10.1016/J.UFUG.2023.128083.
Gao X, Zhao B, Chen Z, Song W, Li Z, Wang X, 2023. The Impact of Urbanization on Tree Growth and Xylem Anatomical Characteristics. Biology 12(11): 1373. https://doi.org/10.3390/BIOLOGY12111373.
Gilman EF, Watson DG, 1994. Populus nigra “Italica” Lombardy Poplar Fact Sheet ST-501. Environmental Horticulture Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida, Florida, USA, 3 p. Available online: https://www.cabidigitallibrary.org/doi/abs/10.1079/cabicompendium.43535 (12 December 2025).
Google earth pro (Desktop), 2025. Belgrade Imagery from 09/2018.
Haarder S, Liston A, 2018. From Dusty Drawers to Verdant Woodlands: New Records of Three Leaf-Mining Sawflies (Hymenoptera: Tenthredinidae) from Northern Europe, with Particular Reference to the Danish Fauna. Entomologiske meddelelser 86 (1-2): 57–73.
Hałaj R, Osiadacz B, 2013. European gall-forming Pemphigus (Aphidoidea: Eriosomatidae). Zool Anz 252(4): 417–423. https://doi.org/10.1016/j.jcz.2013.04.002.
Hammer DAT, Ryan PD, Hammer Ø, Harper DAT, 2001. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontologia Electronica 4: 9.
Helen Jarzebski MP, Gasparatos A, 2019. Land use change, carbon stocks and tree species diversity in green spaces of a secondary city in Myanmar, Pyin Oo Lwin. PLoS One 11: 14. https://doi.org/10.1371/journal.pone.0225331.
Hering EM, 1957. Bestimmungstabellen der Blattminen von Europa einschliesslich des Mittelmeerbeckens und der Kanarishen Inseln, Band I, II und III. Dr W. Junk, Hague, Netherlands, 1395 p. https://doi.org/10.1007/978-94-010-3702-0.
Hering EM, 1951. Biology of the Leaf Miners. Dr W. Junk, Hague, Netherlands, 420 p.
Horák J, Marković Č, Dobrosavljević J, Rada P, Mladenović S, Kohutka A, Míkovcová A, Pech P, Drábek O, Tejnecký V, 2024. Influence of forest structure and soil chemistry on ants, land snails, and beetles in Balkan floodplain forests: Examining species richness and habitat preferences. J Soil Water Conserv 79(6): 303–306. https://doi.org/10.2489/jswc.2024.00045.
Jones EL, Leather SR, 2012. Invertebrates in urban areas: A review. Eur J Entomol 109(4): 463–478. https://doi.org/10.14411/eje.2012.060.
Knuff AK, Staab M, Frey J, Dormann CF, Asbeck T, Klein AM, 2020. Insect abundance in managed forests benefits from multi-layered vegetation. Basic Appl Ecol 48: 124–135. https://doi.org/10.1016/J.BAAE.2020.09.002.
Korányi D, Markó V, 2022. Host plant identity and condition shape phytophagous insect communities on urban maple (Acer spp.) trees. Arthropod Plant Interact 16: 129–143. https://doi.org/10.1007/s11829-022-09887-z.
Kot I, Kmieć K, 2020. Poplar Tree Response to Feeding by the Petiole Gall Aphid Pemphigus spyrothecae Pass. Insects 11(5): 282. https://doi.org/10.3390/insects11050282.
Langmaid L, 2019. British leafminers. Available online: http://www.leafmines.co.uk./ (12 December 2025).
Laštůvka A, Zdeněk L, Liška J, Šumpich J, 2018. Motýli a housenky střední Evropy V., Drobní motýli I. Academia, Praha, Prague, Czech Republic, 536 p.
Lefèvre F, Barsoum N, Heinze B, Kajba D, Rotach P, de Vries SMG, Turok J, 2001. EUFORGEN Technical Bullettin: In situ conservation of Populus nigra. International Plant Genetic Resources Institute, Rome, Italy. Available online: https://www.euforgen.org/uploads/tx_news/741_In_situ_conservation_of_Populus_nigra.pdf (12 December 2025).
Lepiforum, 2019. Lepiforum. Available online: https://www.lepiforum.org/ (12 December 2025).
Levei L, Cadar O, Babalau-Fuss V, Kovacs E, Torok AI, Levei EA, Ozunu A, 2021. Use of Black Poplar Leaves for the Biomonitoring of Air Pollution in an Urban Agglomeration. Plants 10(3): 548. https://doi.org/10.3390/PLANTS10030548.
Liston A, Mutanen M, Viitasaari M, 2019. On the taxonomy of Heterarthrus (Hymenoptera, Tenthredinidae), with a review of the West Palaearctic species. J Hymenopt Res 72: 83–126. https://doi.org/10.3897/jhr.72.39339.
Łukaszkiewicz J, Długoński A, Fortuna-Antoszkiewicz B, Fialová J, 2024. The Ecological Potential of Poplars (Populus L.) for City Tree Planting and Management: A Preliminary Study of Central Poland (Warsaw) and Silesia (Chorzów). Land 13(5): 593. https://doi.org/10.3390/LAND13050593.
Magurran AE, 2004. Measuring Biological Diversity. Blackwell Publishing, Oxford, UK, 256 p.
Marković Č, Dobrosavljević J, Milanović S, 2021a. Factors Influencing the Oak Lace Bug (Hemiptera: Tingidae) Behavior on Oaks: Feeding Preference Does not Mean Better Performance? J Econ Entomol 114(5): 2051–2059. https://doi.org/10.1093/jee/toab148.
Marković Č, Dobrosavljević J, Vujičić P, Cebeci HH, 2021b. Impact of regeneration by shelterwood cutting on the pedunculate oak (Quercus robur) leaf mining insect community. Biologia (Bratisl) 76: 1197–1203. https://doi.org/10.2478/s11756-020-00631-7.
Marković Č, Kanjevac B, Perišić U, Dobrosavljević J, 2024. The effect of the oak powdery mildew, oak lace bug, and other foliofagous insects on the growth of young pedunculate oak trees. Front For Glob Change 6: 1297560. https://doi.org/10.3389/ffgc.2023.1297560.
Marković Č, Stojanović A, Dobrosavljević J, 2018. Diversity and abundance of coccinellids (Coleoptera: Coccinellidae) on trees in parks and tree rows of Belgrade. Biologia (Bratisl) 73: 857–865. https://doi.org/10.2478/s11756-018-0087-5.
Md Meftaul I, Venkateswarlu K, Dharmarajan R, Annamalai P, Megharaj M, 2020. Pesticides in the urban environment: A potential threat that knocks at the door. Sci Total Environ 711: 134612. https://doi.org/10.1016/j.scitotenv.2019.134612.
Meineke EK, Dunn RR, Sexton JO, Frank SD, 2013. Urban Warming Drives Insect Pest Abundance on Street Trees. PLOS ONE 8: e59687. https://doi.org/10.1371/journal.pone.0059687.
Milanović S, Milenković I, Dobrosavljević J, Popović M, Solla A, Tomšovský M, Jankovský L, Tomšovsky M, Jankovský L, 2020. Growth Rates of Lymantria dispar Larvae and Quercus robur Seedlings at Elevated CO2 Concentration and Phytophthora plurivora Infection. Forests 11(10): 1059. https://doi.org/10.3390/f11101059.
Milanović SD, Simović N, Dobrosavljević J, Milenković ILj, Branković Z, Ćirkovi, J, Radojković A, Perać S, Jovanović J, Tadić V, Žugić A, Branković G, 2025. Bioactivity of the Tree of Heaven Leaf Extracts Incorporated into Biopolymer Matrix Against Spongy Moth Larvae. Forests 16(2): 375. https://doi.org/10.3390/f16020375.
Minić R, Josipović M, Spirić VT, Gavrović-Jankulović M, Popadić AP, Prokopijević I, Ljubičić A, Stamenković D, Burazer L, 2020. Impact of tree pollen distribution on allergic diseases in Serbia: Evidence of implementation of allergen immunotherapy to Betula verrucosa. Medicina 56(2): 59. https://doi.org/10.3390/MEDICINA56020059.
Moir ML, Hughes L, Vesk PA, Leng MC, 2014. Which host-dependent insects are most prone to coextinction under changed climates? Ecol Evol 4: 1295–1312. https://doi.org/10.1002/ece3.1021.
Ojija F, Mng’ong’o M, Aloo BN, Mayengo G, Helikumi M, 2025. Effect of global climate change on insect populations, distribution, and its dynamics. J Asia Pac Entomol 28(3): 102442. https://doi.org/10.1016/J.ASPEN.2025.102442.
Oke TR, 1982. The energetic basis of the urban heat island. Q J R Meteorol Soc, 108, 1–24. https://doi.org/10.1002/qj.49710845502.
Parmesan C, 2006. Ecological and evolutionary responses to recent climate change. Annu Rev Ecol Evol Syst 37: 637–669. https://doi.org/10.1146/annurev.ecolsys.37.091305.110100.
Peralta G, Fenoglio MS, Salvo A, 2011. Physical barriers and corridors in urban habitats affect colonisation and parasitism rates of a specialist leaf miner. Ecol Entomol 36, 673–679. https://doi.org/10.1111/j.1365-2311.2011.01316.x.
Percival GC, 2023. Heat tolerance of urban trees − A review. Urban For Urban Green 86: 128021. https://doi.org/10.1016/J.UFUG.2023.128021.
Pincebourde S, Woods HA. 2012. Climate uncertainty on leaf surfaces: The biophysics of leaf microclimates and their consequences for leaf-dwelling organisms. Funct Ecol 26: 844–853. https://doi.org/10.1111/j.1365-2435.2012.02013.x.
Ramsar Convention Secretariat, 2016. Ramsar handbooks for the wise use of wetlands, The fourth strategic plan 2016-2024, 5th edition, vol. 2. ed. Ramsar Convention Secretariat, Gland, Switzerland, 51 p.
Raupp MJ, Shrewsbury PM, Herms DA, 2010. Ecology of Herbivorous Arthropods in Urban Landscapes. Annu Rev Entomol 55: 19–38. https://doi.org/10.1146/annurev-ento-112408-085351.
Rickman JK, Connor EF, 2003. The effect of urbanization on the quality of remnant habitats for leaf-mining Lepidoptera on Quercus agrifolia. Ecography 26, 777–787. https://doi.org/10.1111/j.0906-7590.2003.03345.x.
Robinet C, Roques A, 2010. Direct impacts of recent climate warming on insect populations. Integr Zool, 5: 132–142. https://doi.org/10.1111/j.1749-4877.2010.00196.x.
Rodrıguez-Castañeda G, MacVean C, Cardona C, Hof AR, 2017. What limits the distribution of Liriomyza huidobrensis and its congener Liriomyza sativae in their native niche: When temperature and competition affect species’ distribution range in Guatemala. J Insect Sci 17(4): 88. https://doi.org/10.1093/JISESA/IEX059.
Sallé A, Cours J, Le Souchu E, Lopez-Vaamonde C, Pincebourde S, Bouget C, 2021. Climate Change Alters Temperate Forest Canopies and Indirectly Reshapes Arthropod Communities. Front For Glob Change 4: 710854. https://doi.org/10.3389/ffgc.2021.710854.
Schneider CA, Rasband WS, Eliceiri KW, 2012. NIH Image to ImageJ: 25 years of image analysis. Nat Methods 9(7): 671–675. https://doi.org/10.1038/NMETH.2089.
Schowalter TD, 2006. Insect ecology, An Ecosystem Approach, 2nd ed. Elsevier Inc, Burlington, USA, 576 p.
Schowalter TD, 1995. Canopy invertebrate community response to disturbance and consequences of herbivory in temperate and tropical forests. Selbyana 16(1): 41–48.
Shams MH, Assarehzadegan MA, Eskandari N, Masjedi M, Kheirandish F, Ghasemi R, Ganjalikhani hakemi M, Varzi AM, Safari M, Sohrabi SM, Abdoli Sereshki H, 2021. Molecular and immunochemical characterization of Pop n 2: A new allergen of Populus nigra pollen. Clin Exp Allergy 51: 1613–1623. https://doi.org/10.1111/CEA.13789.
Shannon CE, 1948. A Mathematical Theory of Communication. Bell Syst Tech J 27(3), 379–423. https://doi.org/10.1002/j.1538-7305.1948.tb01338.x.
Šiler B, Skorić M, Mišić D, Kovačević B, Jelić M, Patenković A, Kurbalija Novičić Z, 2014. Variability of European Black Poplar (Populus nigra L.) in the Danube Basin. Public Enterprise “Vojvodinasume”, Petrovaradin, Novi Sad, Serbia, 126 p.
Sinclair JS, Buchner D, Gessner MO, Müller J, Pauls SU, Stoll S, Welti EAR, Bässler C, Buse J, Dziock F, Enss J, Hörren T, Künast R, Li Y, Marten A, Morkel C, Richter R, Seibold S, Sorg M, Twietmeyer S, Weis D, Weisser W, Wiggering B, Wilmking M, Zotz G, Frenzel M, Leese F, Haase P, 2025. Effects of land cover and protected areas on flying insect diversity. Conserv Biol 39: e14425. https://doi.org/10.1111/COBI.14425.
Southwood RTE, Wint WGR, Kennedy CEJ, Greenwood SR, 2004. Seasonality, abundance, species richness and specificity of the phytophagous guild of insects on oak (Quercus) canopies. Eur J Entomol 101(1): 43–50. https://doi.org/10.14411/eje.2004.011.
Southwood TRE, 1961. The Number of Species of Insect Associated with Various Trees. J Anim Ecol 30(1): 1–8. https://doi.org/10.2307/2109.
StatSoft, Inc., 2014. STATISTICA (data analysis software system).
Sunil V, Majeed W, Chowdhury S, Riaz A, Shakoori FR, Tahir M, Dubey VK, 2023. Insect Population Dynamics and Climate Change. In: Ali H, Hou Y, Tahir MB (Eds.) Climate Change and Insect Biodiversity: Challenges and Implications CRC Press, Boca Raton, Florida, USA, 121–146.. https://doi.org/10.1201/9781003382089-8.
Tabassum S, Manea A, Leishman MR, 2024. Limiting the impact of insect pests on urban trees under climate change. Urban For Urban Green 94: 128246. https://doi.org/10.1016/J.UFUG.2024.128246.
Valdés‐Correcher E, Popova A, Galmán A, Prinzing A, Selikhovkin AV, Howe, AG, Mrazova A, Dulaurent A, Hampe A, Tack AJM, Bouget C, Lupaștean D, Harvey D, Musolin DL, Lövei GL, Centenaro G, Halder I, Van Hagge, J, Dobrosavljević J, Pitkänen J, Koricheva J, Sam K, Barbaro L, Branco M, Ferrante M, Faticov M, Tahadlová M, Gossner M, Cauchoix M, Bogdziewicz M, Duduman M, Kozlov MV, Bjoern MC, Mamaev NA, Fernandez‐Conradi P, Thomas RL, Wetherbee R, Green S, Milanović S, Moreira X, Mellerin Y, Kadiri Y, Castagneyrol B, 2022. Herbivory on the pedunculate oak along an urbanization gradient in Europe: Effects of impervious surface, local tree cover, and insect feeding guild. Ecol Evol 12: e8709. https://doi.org/10.1002/ece3.8709.
Vanden Broeck A, 2003. EUFORGEN Technical Guidelines for genetic conservation and use for European black poplar (Populus nigra). International Plant Genetic Resources Institute, Rome, Italy, 6 p.
Xie C, Chen S, Liu D, Jim CY, 2024. Unveiling the complex networks of urban tree diversity research: A global perspective. Ecol Evol 14: e11630. https://doi.org/10.1002/ece3.11630.
© 2026 by the author(s). License: Croatian Forest Research Institute. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution 4.0 International License (CC BY 4.0).