
SEEFOR 15(1): 21-28
Article ID: 2406
DOI: https://doi.org/10.15177/seefor.24-06
ORIGINAL SCIENTIFIC PAPER
Genetic Variation in Provenance-Progeny Trials of Cupre-ssus sempervirens in Türkiye
Alper Ahmet Özbey1,*, Mehmet Çalikoğlu1, Rumi Sabuncu2, Zeynep Gülçin Altun3
(1) Southwest Anatolia Forest Research Institute, Antalya, Türkiye;
(2) Regional Directorate of Forestry (retired), Antalya, Türkiye;
(3) Aegean Forest Research Institute (retired), İzmir, Türkiye
Citation: Özbey AA, Çalikoğlu M, Sabuncu R, Altun ZG, 2024. Genetic Variation in Provenance-Progeny Trials of Cupressus sempervirens in Türkiye. South-east Eur for 15(1): 21-28. https://doi.org/10.15177/seefor.24-06.
Received: 26 Jan 2024; Accepted: 11 Mar 2024; Published online: 10 Apr 2024
Cited by: Google Scholar
Abstract
In this study, the genetic and phenotypic relations among tree height, diameter at breast height and survival at 18 years of age were investigated in Mediterranean cypress (Cupressus sempervirens L. var. horizontalis (Mill.) Gordon) provenance-progeny trials established in two locations in southern Türkiye. It was found that the variation among families within one provenance was higher than among different provenances. At the joint analysis, the narrow-sense heritability (hi2) was 0.36 for height, and 0.24 for diameter. Family mean heritability (hf2) was 0.78 for height, while it was 0.53 for diameter. Diameter showed higher (13.98%) genetic diversity than height (12.71%). Genetic correlation between traits was high (0.84). According to the breeding values of the families estimated by the BLUP method, when the best 50 families by two traits are selected separately, 12.5% genetic gain in terms of height and 13.2% in terms of diameter can be achieved. In the selection of the best 30 families, the estimated genetic gain calculated was 15.2% for height and 16.3% for diameter.
Keywords: cypress; heritability; genetic gain; selection; genetic diversity
INTRODUCTION
Cupressus sempervirens L. var. horizontalis (Mill.) Gordon is a variety of Mediterranean cypress (Cupressus sempervirens L.), which is the only cypress species naturally distributed in Türkiye (Anşin and Özkan 1997). The natural distribution of the Mediterranean cypress is fragmented and discontinuous, extending from the southwestern Mediterranean basin to the Caucasus and southwestern Iran in the east, at altitudes ranging from the sea level (Crete Island) to 2000 m (Türkiye). The Mediterranean cypress is drought-resistant and capable of thriving in areas with an annual precipitation of up to 200 mm (Caudullo and de Rigo 2016).
The natural distribution of Mediterranean cypress in Türkiye is 4,796.9 ha (OGM 2021). Despite its small-scale natural distribution, the species has an important role in forestry studies in Türkiye due to the following reasons:
It may form mixtures with other species that can be used to provide species and product diversity in suitable habitats in potential Turkish red pine (Pinus brutia Ten.) afforestation areas (Sabuncu and Çalışkan 2008), especially considering its fast growth in juvenile age and useful wood (Saatçioğlu 1976).
Its branched stem and dense coniferous top structure make it an effective barrier against wind, dust, noise, and unsightly views. (Caudullo and de Rigo 2016).
Research in pharmacology has revealed that this aromatic plant has crucial medicinal properties that protect against bacteria, fungi, and viruses, repel parasites and insects, and increase resistance to cancer due to its chemical components (Al-Snafi 2016).
It is a tree species with high landscape value (Farjon and Filer 2013). Not only due to its body form, but also because it functions as a solitary object in the urban landscape. On the other hand, with its vertical silhouette, it cuts and balances the static horizontal lines in the landscape. In addition, thanks to its dark green color tones, it creates a visual contrast in the form of scattered spots in the light green and brightly colored Mediterranean vegetation (Çalıkoğlu et al. 2010).
It is one of the tree species used by the General Directorate of Forestry of Türkiye within the scope of the Rehabilitation of Burned Areas and the Establishment of Forest with Fire Resistant Species Projects (OGM 2010).
Due to the reasons mentioned above, silviculture and arboriculture of Mediterranean cypress are gaining importance day by day. In this respect, it is important to determine quality seeds and saplings, best seed source, breeding potential and protection of natural populations. Additionally, it is a necessity to protect Mediterranean cypress’s natural gene resources and develop appropriate policies against the risks posed by various biotic and abiotic factors. In the early studies on the Mediterranean cypress in the nursery, Sabuncu (2004) reported high genetic diversity and heritability for seedling height within the population. Uslu and Bilir (2020) stated that the species outperformed pyramidal cypress in seedling height and diameter at root collar. Regarding the knowledge and general information on the cypress, some prominent studies include: Farahmand (2020), on genetic-breeding studies; Korol et al. (1997), Capuana et al. (2000), Papageorgiou et al. (2005), Gallis et al. (2007), Al-Hawija et al. (2014), Nocetti et al. (2017), Bagnoli et al. (2020), and Ismael et al. (2021).
In this study, the genetic variation, heritability and genetic correlation, genetic gain of height and diameter at breast height, as well as the distribution of genetic variance between and within populations in provenance-progeny trials of Mediterranean cypress were studied.
MATERIALS AND METHODS
Materials
Material of the study consisted of 204 Mediterranean cypress families from 14 provenances (populations). Trees belonging to these provenances were grown from seeds collected from the provenances in the natural distribution area of the Mediterranean cypress in 1999 (Table 1). To keep the kinship relations between the saplings at a minimum level, the distance among the selected trees was attentively 100 meters or less. Seeds collected from each family were planted separately in Enso-type trays in Denizli Forest Nursery Directorate in 2000 (Sabuncu 2004).
![]() Table 1. Details of the provenances and families.
Table 1. Details of the provenances and families.
Trials were established with 1+0 seedlings produced in the 2000-2001 planting season in Antalya and Fethiye regions (Çalıkoğlu et al. 2010) (Table 2). The trials were situated in the same Mediterranean bioclimate layer, and the climatic conditions were similar to each other. Trials were established with 17 provenances. In the preliminary evaluations, statistically significant difference was not observed between the two provenances selected from Fethiye region and the three origins selected from Kemer region for both height and diameter. Therefore, Fethiye and Kemer were accepted as provenances for further analysis. Completely randomized block design with 4-row plot configuration was used in all trials with 5 blocks.
![]() Table 2. Details of the trials.
Table 2. Details of the trials.
Methods
At the end of the 2018 growing season (18th year of the trials), the height (cm) and diameter at breast height (mm) of the trees in both trials were measured and the survival rate was observed. The provenances were compared by multiple analyses of variance (MANOVA). Families’ genetic parameters were estimated based on the mixed model. BLUP (Best Linear Unbiased Prediction) method was used to estimate the breeding values of families. Analysis of variance in the survival rate was determined for the averages of families in a block. Provenances were grouped by Duncan’s multiple range test (Duncan 1955) based on the results of analyses of variance.
The following model was used in the analysis of each trial:
Yijkl = µ + Bi + Pj + Fk(j) + BPij + BFik(j) + eijkl (1)
where Yijkl: observation from lth tree of the kth family in the jth provenance in the ith block, µ: overall mean, Bi: block effect, Pj: provenance effect, Fk(j): family within provenance effect, BPij: block-provenance effect, BFik(j): block-family effect, eijkl: random error.
Yijkmn = μ + Si + Bj(i) + Pk + Fm(k) + SPik + SFim(k) + BPj(i)k + BFjk(im) + eijkmn (2)
where Yijkmn: observation from nth tree of the mth family of the kth provenance in the jth block in the ith provenance, Si: trial effect, Bj(i): block within trial effect, Pk: provenance effect, Fm(k): family within provenance effect, SPjk: trial-provenance effect, SFim(k): trial-family effect, BPj(i)k: block-provenance effect, BFjk(im): block-family effect. The analyses were carried out on 196 families that had common traits in both trials.
If there are significant differences between provenances in provenance-progeny trials, this is also reflected in heritability. The population or provenance effect should be freed from genetic variance (Williams et al. 2002). For this reason, while calculating the variance components and genetic parameters at joint analysis, the provenance effect was eliminated by taking the family variance.
Narrow-sense heritability (hi2) was estimated as (3) (Becker 1984, Coterril 1987):
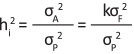 (3)
(3)
where σA2 is the additive genetic variance, σF2 is the genetic variance of families, σP2 is the phenotypic variance, k is the covariance coefficient between half-sibs.
Family mean heritability (h2f) was estimated as (4) (Liu et al. 2013):
h2f = σ2f / (σ2f + σ2sf / s + σ2fb / sb + σ2e / sbn) (4)
where σ2f is the family variance, σ2fb is the family-block variance, σ2sf is the trial-family variance, σ2e is the error variance, s is the trial number, b is the block number, and n is the harmonic mean of the number of trees per family.
Genetic correlation (rg) between tree height and diameter at breast height were estimated as (5) (Falconer 1996):
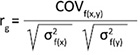 (5)
(5)
where COVf(x,y) is the genetic covariance between traits x and y, and σ2f(x) and σ2f(y) are the additive genetic variances of traits x and y, respectively.
The Delta Method was used to calculate the standard errors of the heritabilities and genetic correlation (Lynch and Walsh 1997). Type B genetic correlations between trials were determined by the following equation (Burdon 1977):
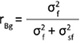 (6)
(6)
Coefficient of variation of genetic (CVg) was estimated based on genetic σ2f variance, and mean (x_) of the traits according to the following equation (Sun 1980):
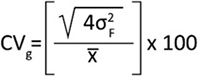 (7)
(7)
Genetic gain (ΔG (%)) was estimated as:
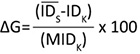 (8)
(8)
where IDS is the average of breeding value of the highest 30 families, IDK is the breeding value of the control material, and MIDK is the absolute breeding value of the control material.
RESULTS
Some descriptive statistics of the measured traits are given in Table 3. Survivals in the Antalya and Fethiye trials were 84% and 91%, respectively. Averages of height were 748.04 cm in Antalya and 627.82 cm in Fethiye. Means of diameter were 103.54 mm in Antalya and 93.58 mm in Fethiye (Table 3).
![]() Table 3. Descriptive statistics of the measured traits.
Table 3. Descriptive statistics of the measured traits.
Antalya Trial
The differences between provenances were not significant (P>0.05) for both traits (Table 4).
![]() Table 4. Results of analysis of variance for Antalya.
Table 4. Results of analysis of variance for Antalya.
Significant differences (P<0.05) were found between provenances for survival rate (Table 5) and the provenances were grouped (Table 6).
![]() Table 5. Results of analysis of variance for survival rate between provenances.
Table 5. Results of analysis of variance for survival rate between provenances.
![]() Table 6. Homogeneous groups in terms of survival rate between provenances.
Table 6. Homogeneous groups in terms of survival rate between provenances.
The family effect on survival rate was significant (P=.0011) (Table 7). The lower limit of the best family group had a 65% survival rate. Families under this limit were 1324, 3401, 1102, 2148, 3312, 1405 and 3101.
![]() Table 7. Results of analysis of variance for survival rate between families.
Table 7. Results of analysis of variance for survival rate between families.
Fethiye Trial
The differences between provenances were significant (P<0.05) for both traits (Table 8) and the provenances were grouped (Table 9).
![]() Table 8. Results of analysis of variance for Fethiye.
Table 8. Results of analysis of variance for Fethiye.
![]() Table 9. Homogeneous groups in terms of height and diameter between provenances.
Table 9. Homogeneous groups in terms of height and diameter between provenances.
Significant differences (P<0.05) were found between provenances for survival rate (Table 10) and the provenances were grouped (Table 11).
![]() Table 10. Results of analysis of variance for survival rate between provenances.
Table 10. Results of analysis of variance for survival rate between provenances.
![]() Table 11. Homogeneous groups in terms of survival rate between provenances.
Table 11. Homogeneous groups in terms of survival rate between provenances.
Family effect on the survival rate in the Fethiye trial area was also significant (Table 12). The lower limit of the best family group in this trial area had an 80% survival rate. Families under this limit were 3301, 2402, 3102, 1203, 1324, 2303, 1408, 1102, 3101 and 3111.
![]() Table 12. Results of analysis of variance for survival rate between families.
Table 12. Results of analysis of variance for survival rate between families.
Joint Analysis
In joint analysis of variance for trials, while provenance effect for height was significant, provenance effect for diameter was not significant (Table 13). Provenances were grouped according to height (Table 14).
![]() Table 13. Joint analysis of variance for trials.
Table 13. Joint analysis of variance for trials.
![]() Table 14. Homogeneous groups in terms of height between provenances.
Table 14. Homogeneous groups in terms of height between provenances.
The variance components of the joint analysis of the trials for both traits are given in Table 15. The ratio of the variance component for provenances was estimated as 0.92% for diameter and 2.38% for height.
Genetic Parameters
Some genetic and phenotypic parameters of traits were calculated in joint analysis (Table 16). Narrow-sense (hi²) and family mean heritability (hf²) for height were bigger than for the diameter. Coefficient of genetic variation (CVg) was estimated as 13.98 for diameter and 12.71 for height. The correlation between trials (Corr bG) was estimated as 0.83 for height and 0.68 for diameter. High genetic and phenotypic correlations were observed between height and diameter.
![]() Table 16. Some genetic and phenotypic parameters in joint analysis.
Table 16. Some genetic and phenotypic parameters in joint analysis.
In the joint analysis, it was estimated that a 12.5% genetic gain in height and a 13.2% genetic gain in diameter could be achieved by selecting the best 50 families from 196 families. With the selection of the best 30 families, these rates increased to 15.2% and 16.3%, respectively. 37 of the best 50 families in terms of diameter were also included in the best 50 families in terms of height. The participation rate of provenances in the top 50 families for height ranged from 2% (Datça and Kaş) to 28% (Kemer).
DISCUSSION
According to the 18th year assessment on field evaluations of Mediterranean cypress provenance-progeny trials, Antalya has shown better performance for both height and diameter growth. In the 8th year assessment of the trials, it was determined that the Fethiye trial showed better growth and higher survival rate. It was stated that this situation may have arisen from the deep field soil in the trial (Çalıkoğlu et al. 2010). However, in the 18th year assessment, Antalya trial showed 16% improvement in height. Lower growth in the Fethiye trial was attributed to a solid gravel layer located approximately 70 cm below the surface.
It was observed that the variation between families within the provenances was higher than the variation between the provenances for height and diameter in Mediterranean cypress (Table 15). This result illustrates that selections within the provenances will be more important than the selection of provenance in a breeding program to be applied for growth of Mediterranean cypress.
On the other hand, in Mediterranean cypress, in-provenance selections may result in higher genetic gain as well as in a breeding population with higher genetic variation. For instance, if only 30 families with the best breeding value were selected in Köprülü Canyon (21), the genetic gain for height growth would be 7%. However, regardless of provenance, the genetic gain in height with the selection of the top 30 families in the trials was estimated as 15.2%. Moreover, these 30 families belong to 9 different provenances.
A comparison can be made for narrow-sense heritability of 18-year-old trees’ height and diameter traits in Mediterranean cypress with estimations from a recent Turkish red pine progeny trial.
In the 16th year assessment of low elevation breeding zone (0-400 m) of the Aegean Region Turkish red pine progeny trials, narrow-sense heritability of height and diameter was estimated as 0.27 and 0.28, respectively (Özyalçın et al. 2016). These values were found to be 0.36 and 0.24 in Mediterranean cypress, respectively. Family mean heritability of height and diameter in Turkish red pine was estimated as 0.49 and 0.59, respectively. In Mediterranean cypress these values were estimated as 0.78 and 0.53, respectively. In Turkish red pine, CVg varied between 8% and 13% in height and 10% and 18% in diameter, depending on the trial areas. In the Mediterranean cypress, it was estimated as 12.71% for height and 13.98% for diameter.
According to the 18th year assessment of the Mediterranean cypress trials, the genetic gains to be obtained by selecting 50 or 30 families with the best breeding value in terms of height and diameter were also estimated. A population of 50 families that will form the production population (seed plantation) will form a wide-ranging basis for future thinning. A production population (seed plantation) of 50 families will provide a wide-ranging basis for future thinning. A population of 30 families will yield a higher genetic gain. As a matter of fact, this number is accepted as a number that will not significantly reduce genetic variation (Namkoong et al. 1988, Varela and Eriksson 1995).
The genetic gains to be achieved at the age of 18 in terms of height in Mediterranean cypress were estimated as follows: 12.5% in the selection of 50 families, 15.2% in the selection of 30 families with the best breeding value in terms of height. In terms of diameter, these values were: 13.2% in the selection of the best 50 families and 16.3% in the selection of the best 30 families.
High genetic correlation (r=0.84) was estimated between height and diameter in Mediterranean cypress. This means that the selection to be made for any trait in Mediterranean cypress will not significantly change the genetic gain (Zobel and Talbert 1984).
Diameter is a parameter that affects volume more than height, and its measurement is easier and more practical. If the genetic relationship between the two traits persists at this level in the mature age, it would be more beneficial to make the selection according to the diameter.
Type B genetic correlation (r=0.83) between trials in terms of height was high. In terms of diameter, this value was estimated at the accepted limit value (0.68) for sub-zoning. In the 8th year assessment, it was calculated for diameter as 0.85 and for height as 0.81 (Çalıkoğlu et al. 2010).
The survival rate was significantly affected by family in both trials. During the selection of breeding plantations, it would be useful to pay attention to families that are not in the best group in terms of survival rate.
CONCLUSIONS
The current family set of Mediterranean cypress selected within the scope of this study should be considered as the main breeding population of this species. The basic genetic parameters of the species have been revealed in this study. However, the increasing demand for Mediterranean cypress saplings (border tree, mixture type in Turkish red pine plantations) reveals the necessity of establishing a seed garden or a seed plantation. By deciding on a selection intensity that they deem appropriate, tree breeders will be able to benefit from the superior families determined by this study. During the establishment phase, it is recommended to begin with 40-50 families that have the best breeding value in terms of diameter. In the following years, it is advised to reduce the number of families to 20-30, as revealed in this study.
Author Contributions
RS and ZGA established the trials, AAÖ and ZGA collect the data, AAÖ and MÇ performed statistical analyses. AAÖ and MÇ drafted the manuscript, RS and ZGA improved the manuscript draft. All authors discussed the results and contributed to the final manuscript.
Funding
This research was supported by Turkish General Directorate of Forestry for data collection.
Conflicts of Interest
The authors declare no conflict of interest.
REFERENCES
Al-Hawija BN, Wagner V, Hensen I, 2014. Genetic comparison between natural and planted populations of Pinus brutia and Cupressus sempervirens in Syria. Turkish Journal of Agriculture and Forestry 38(2): 267-280. https://doi.org/10.3906/tar-1211-24.
Al-Snafi E, 2016. Medical importance of Cupressus sempervirens- A Review. IOSR Journal of Pharmacy 6 (6): 66-76.
Anşin R, Özkan ZC, 1997. Tohumlu Bitkiler (Spermathophyta): Odunsu Taksonlar. Karadeniz Technical University Faculty of Forestry Trabzon 167(19): 306–311.
Bagnoli F, Della Rocca G, Spanu I, Fineschi S, Vendramin GG, 2020. The origin of Afro-Mediterranean cypresses: Evidence from genetic analysis. Perspectives in Plant Ecology, Evolution and Systematics 46: 125-564. https://doi.org/10.1016/j.ppees.2020.125564.
Becker WA, 1984. Manual of Quantitative Genetics. 4th edition, Academic Enterprises, Washington, USA, 190p.
Burdon RD, 1977. Genetic correlation as a concept for studying genotype-environment interaction in forest tree breeding. Silvae Genetica 26(5-6): 168-175.
Capuana M, Giovannelli A, Giannini R, 2000. Factor influencing rooting in cutting propagation of Cypress (Cupressus sempervirens L.). Silvae Genetica 49(6): 277-281.
Caudullo G, De Rigo D, 2016. Cupressus sempervirens in Europe: distribution, habitat, usage and threats. In: San-Miguel-Ayanz J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A. (Eds.), European Atlas of Forest Tree Species. Publ. Off. EU, Luxembourg, 88-89.
Coterril PP, 1987. Shorte note: On estimating heritability according to practical applications. Silvae Genetica 36(1): 46-48.
Çalıkoğlu M, Sabuncu R, Baş MN, Coşgun S, Altun ZG, Çetinay Ş, 2010. Mediterrenean Cypress (Cupressus sempervirens L. var. horizontalis (Mill.) Gordon) provenance-progeny trials (Results of 8th age). Southwest Anatolia Forest Research Institute, Technical Bulletin, 36, Antalya.
Duncan DB, 1955. Multiple range and multiple F tests. Biometrics 11(1): 1-42.
Falconer DS, Mackay FCT, 1996. Introduction to Quantitative Genetics. 4th edition, Longman, Edinburg, Scotland, 464p.
Farahmand H, 2020. The genus Cupressus L. Mythology to biotechnology with emphasis on Mediterranean cypress (Cupressus sempervirens L.). Horticultural Reviews 47: 213-287.
Farjon D, Filer D, 2013. An Atlas of the World’s Conifers: An Analysis of their Distribution, Biogeography, Diversity and Conservation Status. Brill, Leiden, Netherlands.
Gallis AT, Doulis AG, Papageorgieu AC, 2007. Variability of cortex terpene composition in Cupressus sempervirens L. provenances grown in Crete-Greece. Silvae Genetica (56)6: 294-299. https://doi.org/10.1515/sg-2007-0042.
Ismael A, Klápšte J, Stovold GT, Fleet K, Dungey H, 2021. Genetic Variation for Economically Important Traits in Cupressus lusitanica in New Zealand. Frontiers in Plant Science 12:651729. https://doi.org/10.3389/fpls.2021.651729.
Korol L, Kara N, Işık K, Schiller G, 1997. Genetic differentiation among and within natural and planted Cupressus sempervirens L. Eastern Mediterrenean populations. Silvae Genetica 46(2-3): 151-155.
Lynch M, Walsh B, 1997. Genetics and Analyses of Quantitative Traits. Sinauer Ass. Inc. Publ., Sunderland, USA.
Liu QH, Zhou ZC, Fan HH, Liu YR, 2013. Genetic variation and correlation among resin yield, growth, and morphologic traits of Pinus massoniana. Silvae Genetica 62: 38–44. https://doi.org/10.1515/sg-2013-0005.
Namkoong G, Kang HC, Brouard JS, 1988. Tree breeding: principles and strategies: principles and strategies. 1st ed. Springer, New York, USA, 180p.
Nocetti M, Della Rocca G, Berti S, Brunetti M, Di Lonardo V, Pizzo B, Danti R, 2017. Clonal consistency of wood technological properties in canker-resistant Cupressus sempervirens clones at two contrasting sites. Tree genetics & genomes, 13(2): 1-12. https://doi.org/10.1007/s11295-017-1111-6.
Uslu M, Bilir N, 2020. Morphological Seedling Characteristics in Mediterranean and Pyramidal Cypresses. The Journal of Graduate School of Natural and Applied Sciences of Mehmet Akif Ersoy University 11(1): 12-21. https://doi.org/10.29048/makufebed.681763.
OGM, 2010. Rehabilitation of Burned Areas and the Establishment of Forest with Fire-resistant Species Project (YARDOP). Circular letter, 6665, General Directorate of Forestry, Turkey.
OGM, 2021. Official Statistics, Forestry Statistics. Accessed: https://www.ogm.gov.tr/tr/e-kutuphane/resmi-istatistikler. Available online: 17.02.2022.
Özyalçın K, Ezen T, Altun ZG, Ünal C, Alan M, Öztürk H, Şıklar S, Derilgen SI, 2016. Turkish Red Pine (Pinus brutia Ten.) Progeny Trials in Low Elevation Breeding Zone (0-400 m) of Aegean Region (16th Year Results). Forest Tree Seeds and Tree Breeding Research Directorate, Interim Results Report, Ankara, Turkey.
Papageorgiou AC, Finkeldey R, Hattemer HH, Xenopoulos S, 2005. Genetic differences between autochthonous and breeding populations of common cypress (Cupressus sempervirens L.) in Greece. European Journal of Forest Research, 124(2): 119-124. https://doi.org/10.1007/s10342-005-0063-9.
Saatçioğlu F, 1976. Silviculture 1. İstanbul University Faculty of Forestry, İstanbul, Turkey.
Sabuncu, R. 2004. Genetic diversity in Mediterrenean Cypress (Cupressus sempervirens L. var. horizontalis (Mill.) Gordon) populations for seedling growth characteristics. Southwest Anatolia Forest Research Institute, Technical Bulletin 22, Antalya, Turkey.
Sabuncu R, Caliskan S, 2008. Ecology and silviculture of Mediterranean cypress (Cupressus sempervirens L.). Journal of the Faculty of Forestry Istanbul University 58(1):53–72.
Sun O, 1980. Statistical methods. Forest Research Institute, Ankara, Turkey, 376 p.
Varela MC, Eriksson G, 1995. Multipurpose gene conservation in Quercus suber-A Portuguese example. Silvae Genetica 44(1):28-37.
Williams ER, Matheson AC, Harwood CE, 2002. Experimental design and analysis for tree improvement. 2nd ed. Canberra: CSIRO.
Zobel B, Talbert J, 1984. Applied forest tree improvement. John Wiley & Sons, New York, USA, 505p.
© 2024 by the Croatian Forest Research Institute. This is an Open Access paper distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0).